
Chitosan Biostimulants in Vegetable and Medicinal Plant Production
- Entoplast

- 3 days ago
- 8 min read
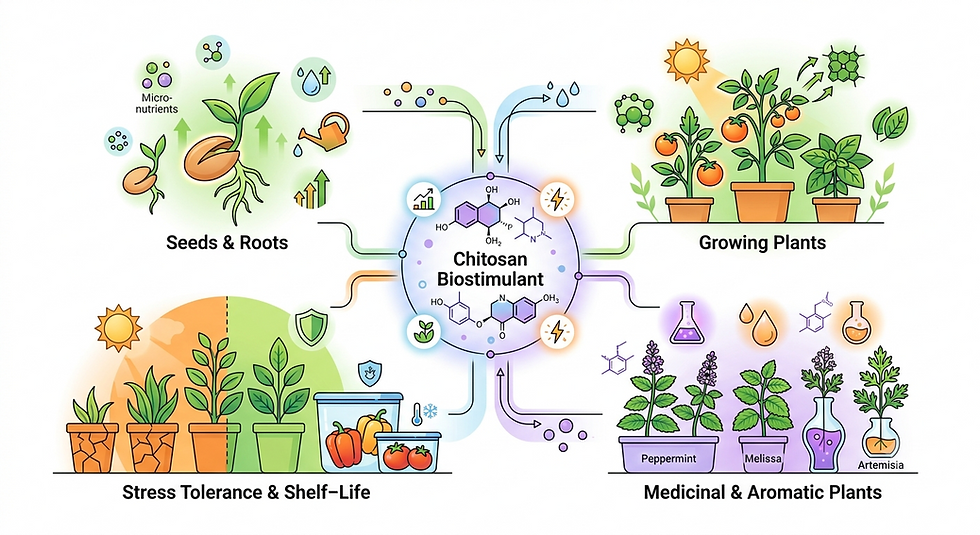
Chitosan‑based biostimulants can genuinely add value in vegetable and medicinal/aromatic crops when they are used as part of a structured programme to support growth, nutrient uptake and stress tolerance, rather than as a stand‑alone “miracle input”. Their performance depends strongly on formulation (molecular weight, degree of deacetylation, bulk vs oligo/nano forms) and on how they are integrated with fertilisers and crop‑protection strategies.
Why chitin and chitosan matter in modern horticulture
Chitosan is a partially deacetylated derivative of chitin, a structural polysaccharide abundant in crustacean shells, fungal cell walls and insect exoskeletons. It is biodegradable, generally non‑toxic and has multidirectional bioactivity: plant growth stimulation, defence elicitation, antimicrobial and antioxidant effects. Reviews in vegetable and medicinal/aromatic crops consistently report that appropriately formulated and dosed chitin/chitosan products can increase biomass and yield, improve nutrient status, enhance tolerance to abiotic stresses, and reduce disease pressure.
Entoplast operates in this space as a supplier of high‑quality chitin and chitosan for horticultural biostimulant and biocontrol formulations, with expertise in controlling molecular weight, deacetylation and purity—parameters that strongly influence agronomic performance. This technical positioning is particularly relevant as growers and formulators look to move towards higher‑value, lower‑impact systems in greenhouse and vertical‑farm production.
What chitosan biostimulants do in horticultural crops
Across vegetables and speciality crops, chitosan and its derivatives act in several overlapping roles:
Growth and yield stimulation: Many studies report improved germination, root development, vegetative growth and fruit/leaf yield after seed coating, substrate amendment or foliar application of chitosan.
Nutrient absorption and metabolism: Chitosan treatments have been shown to enhance photosynthetic pigments, carbon and nitrogen metabolism, and macro/micronutrient uptake, often translating into improved nutrient‑use efficiency.
Abiotic stress tolerance: Under drought, salinity and temperature extremes, chitosan frequently increases chlorophyll content, antioxidant enzyme activity and osmolyte accumulation, helping plants maintain growth and productivity.
Biotic stress and shelf‑life: Chitosan can directly inhibit fungi and bacteria and induce plant defence responses, reducing disease incidence and extending shelf‑life of harvested vegetables and herbs.
Mechanistically, these outcomes reflect chitosan’s ability to induce defence signalling pathways (salicylic acid, jasmonic acid, ethylene), up‑regulate pathogenesis‑related proteins and defence enzymes, and enhance antioxidant systems that limit oxidative damage. Its cationic nature also allows interactions with cell walls, membranes and nutrients, supporting improved nutrient uptake and microbial suppression.
Evidence patterns in vegetable crops

The vegetable‑crop literature is now broad enough to show consistent patterns without pretending that responses are uniform. A 2021 review on vegetable production highlighted that chitin/chitosan biostimulants commonly improved growth, yield and quality in tomato, pepper, cucumber, lettuce, spinach, onion, bean and basil under both optimal and stress conditions.
Key themes include:
Seed treatments (bulk or oligochitosan) often improve germination rate, seedling vigour and early root development in crops such as common bean, cucumber, artichoke and potato.
Early‑stage foliar sprays in lettuce, tomato, spinach and pepper increase leaf area, chlorophylls and carotenoids, and can enhance defensive phytochemicals such as phenolics and flavonoids.
In water‑limited or saline conditions, foliar or root‑zone applications have mitigated yield losses in tomato, onion, okra and basil by sustaining photosynthesis and activating antioxidant defences.
Post‑harvest coatings on fresh‑cut pepper, tomato and other vegetables delay microbial spoilage and quality decline, extending shelf‑life without conventional synthetic preservatives.
At the same time, some trials report neutral or inconsistent effects, particularly when product composition is poorly defined or doses are sub‑optimal. These mixed results underline the need for standardised formulations and careful programme design, rather than assuming a generic “chitosan effect” across all vegetable systems.
Evidence in medicinal and aromatic plants
Medicinal and aromatic plants such as Achillea millefolium, Ocimum basilicum, Melissa officinalis, Mentha piperita, Curcuma longa and Artemisia annua are particularly responsive to inputs that influence secondary metabolite pathways. A recent review on chitosan/chitin‑based biostimulants in medicinal and aromatic plants concluded that low‑to‑moderate doses, applied via foliar sprays or soil amendments, can increase biomass and the concentration of key active compounds, while improving stress tolerance.
Reported outcomes include:
In basil and peppermint, chitosan foliar sprays enhanced essential‑oil yield and modified oil composition towards higher antioxidant and antimicrobial activity, especially under drought stress.
In Achillea, Melissa and other Lamiaceae, chitosan increased total phenols, flavonoids and related antioxidant capacity, supporting both agronomic resilience and phytochemical quality.
In Curcuma and Artemisia, soil or foliar applications have been associated with higher rhizome/leaf biomass and increased accumulation of specific medicinal compounds under biotic and abiotic stress.
Importantly, not all medicinal/aromatic species respond in the same way or at the same dose; some exhibit plateau or even declines in metabolite content at higher chitosan concentrations, reinforcing the need for crop‑specific optimisation.
Mechanisms and formulation choices that matter in practice
Defence elicitation and stress physiology
Mechanistic work in cereals and horticultural species shows that chitosan acts as an elicitor, perceived by plant receptors and triggering defence signalling cascades. These cascades lead to increased activity of enzymes such as peroxidase, catalase, superoxide dismutase and phenylalanine ammonia‑lyase, as well as accumulation of phenolics and other antioxidant metabolites.
Under salinity and other abiotic stresses, chitosan and chitosan nanoparticles have been shown to:
Enhance osmolyte accumulation (e.g. proline, soluble sugars).
Modulate ion homeostasis by affecting Na⁺ compartmentalisation and K⁺ retention.
Increase membrane stability and reduce reactive oxygen species (ROS) production.
These responses underpin observed improvements in plant height, biomass, pigment content and, in some cases, essential‑oil or anthocyanin levels in stress‑exposed horticultural crops.
Root development and nutrient uptake

Metabolite profiling and physiological studies indicate that chitosan can alter carbon and nitrogen metabolism, increase organic acids and sugars, and influence root architecture, resulting in greater nutrient foraging and uptake. Nanochitosan formulations, in particular, have been reported to improve fertiliser efficiency and nutrient content of bulbs in crops like onion.
For agronomists, the takeaway is that chitosan should be thought of as both a stress‑physiology tool and a modulator of root and nutrient dynamics, which is why timing around establishment and early vegetative growth often delivers strong responses.
Molecular weight, deacetylation and particle size
Commercial chitosan products differ markedly in origin, molecular weight (MW), degree of deacetylation (DD), acetylation pattern and any chemical modifications. These parameters affect solubility, charge density, viscosity and biological activity:
Bulk high‑MW chitosan is often less soluble and can become viscous at higher concentrations, with some reports of root growth inhibition when used at non‑optimal doses.
Oligochitosan (shorter chains) tends to be more soluble, with strong elicitor activity at relatively low concentrations and lower risk of physical impediment in the rhizosphere.
Nanochitosan can further increase bioactivity and allow controlled delivery or synergistic combinations with fertilisers and other biostimulants, but raises additional questions around regulatory treatment of nano‑materials and long‑term environmental fate.
Standardisation and transparent specification of MW, DD and formulation type are still works in progress in the commercial market, complicating direct comparison between products and trials. This is an area where technically focused suppliers such as Entoplast, capable of tightening and documenting these parameters, can materially support more reproducible agronomic outcomes.
Designing realistic biostimulant programmes

Rather than seeking universal “recipes”, growers and agronomists should design chitosan programmes around crop stage, target outcome and local constraints, using typical dose ranges as a starting point and adapting based on product labels and field response.
1. Seed coating and substrate amendment (establishment and roots)
Objective: Improve emergence, early vigour and root development, particularly under sub‑optimal moisture or pathogen pressure.
Indicative practice:
Seed coating: bulk or oligochitosan at roughly 0.5–3 g kg⁻¹ seed, often combined with micronutrients or biofertilisers.
Substrate amendment: 0.5–2 kg ha⁻¹ equivalent of chitosan incorporated into nursery media or transplant beds.
Chitosan coatings have reduced seed‑borne fungal contamination and improved germination and root fresh weight in crops such as artichoke, potato and lettuce. When integrating with fertiliser, avoid heavily saline or strongly alkaline carriers that may reduce chitosan solubility and activity.
2. Early vegetative foliar applications (growth and stress priming)
Objective: Enhance canopy development, photosynthetic capacity and stress preparedness in vegetables and herbs.
Indicative practice:
Foliar sprays at 50–200 mg L⁻¹ chitosan equivalent, applied 2–4 times from early vegetative stage to pre‑flowering, with 7–14‑day intervals.
Can be tank‑mixed with conventional fertilisers, seaweed extracts or humic substances, subject to compatibility testing (jar test and small plot trial).
Studies in lettuce, tomato, spinach, basil and pepper show increased leaf area, chlorophylls, phenolic content and, under drought, improved antioxidant status and yield stability under such regimes.
3. Targeted sprays around flowering and metabolite accumulation (medicinal/aromatic crops)
Objective: Support biomass and secondary metabolite production in essential‑oil and medicinal crops.
Indicative practice:
Foliar applications at 50–150 mg L⁻¹ during pre‑flowering and early flowering, with 2–3 sprays timed to known windows of oil or metabolite accumulation.
Optionally combined with micronutrient or elicitor cocktails (e.g. salicylic acid, seaweed extracts), with care not to overload plants with multiple strong signals at once.
Such programmes have increased essential‑oil yield and modified composition in basil and peppermint, and enhanced phenolic profiles in Achillea and Melissa, particularly when plants are under moderate abiotic stress that leaves some “headroom” for stress‑priming benefits.
4. Post‑harvest coatings (quality and shelf‑life)
Objective: Reduce microbial spoilage and maintain texture and visual quality of fresh vegetables and herbs.
Indicative practice:
Dipping or spraying harvested produce with chitosan solutions at 5–20 g L⁻¹, sometimes combined with organic acids or mild sanitising agents.
Application to fresh‑cut pepper, tomato, strawberries and leafy herbs has delayed mould growth, reduced weight loss and maintained marketable quality for several extra days in storage.
Post‑harvest operations must work within food‑contact regulations, and nanochitosan coatings, in particular, may face distinct regulatory scrutiny.
Across all four programme types, the emphasis should be on small‑scale pilot trials with clear metrics (growth, yield, quality, stress indicators, residue/microbiome assessments), rather than immediate broad deployment.
Sourcing, BSF‑derived chitosan and sustainability
Traditionally, commercial chitosan has been sourced from crustacean shells, raising questions about seasonality, supply concentration and potential contamination with heavy metals or allergens. Insects, and black soldier fly (BSF) in particular, are emerging as sustainable alternative sources of chitin and chitosan that align with circular‑economy principles.
BSF production offers several advantages:
Larvae can be reared on a wide range of organic residues, converting low‑value waste into protein, lipids and fertiliser, with chitin‑rich exuviae and cuticles as side streams.
Chitin yields from BSF larvae and pupal exuviae are comparable to crustacean sources, and deacetylated BSF chitosan can achieve degrees of deacetylation and structural properties similar to shrimp chitosan.
Greener extraction processes are being developed to reduce chemical footprints in insect chitosan production, although fully “green” workflows to chitosan are not yet commonplace.
Recent work has shown BSF‑derived chitosan fractions acting as effective plant biostimulants and protectants—for example, improving lettuce biomass and chlorophyll content in hydroponic systems, priming tomato defence against Fusarium, and enhancing Beta vulgaris seed germination and disease resistance. Chitin‑fortified BSF compost has also been used to suppress nematodes and improve potato yield, illustrating how insect‑based inputs can integrate fertility and biocontrol functions.
However, most published horticultural trials still rely on crustacean‑derived materials, and BSF‑specific data in high‑value greenhouse vegetables and medicinal/aromatic crops remain limited. For now, BSF‑derived chitosan should be positioned as a technically viable, sustainable alternative that can be engineered to match the specifications used in existing studies, with R&D and pilot programmes needed to document crop‑ and formulation‑specific performance.
Entoplast’s interest in BSF‑derived chitin/chitosan fits into this emerging landscape: by upcycling insect side streams into well‑characterised polymers, suppliers can help horticultural systems decouple from marine waste and diversify input supply chains.
Regulatory and strategic context
Regulatory treatment of chitosan‑based inputs varies by jurisdiction: in Europe, chitosan has been considered both as a plant biostimulant and as a “basic substance” or biocontrol agent, depending on use pattern and claims. Nanochitosan and heavily modified chitosan conjugates may fall under specific nano‑material or plant‑protection regulations, and any use in edible crops or post‑harvest coatings must comply with food‑contact and residue frameworks.
Strategically, chitosan biostimulants fit with horticulture’s move towards reduced synthetic pesticides, improved product quality and stronger sustainability credentials, especially where they enable lower agrochemical loads while maintaining or improving yield and quality. For agronomy and R&D teams, the practical priorities are to:
Build data on crop‑ and context‑specific responses, including economic assessments.
Clarify interactions with fertilisers, biofertilisers and other biostimulants.
Ensure product positioning and label claims align with local regulatory categories (biostimulant, plant‑protection product, basic substance, etc.).
Entoplast’s role as a technically focused supplier and R&D partner is to connect emerging science on chitosan biostimulants, including BSF‑derived grades, with commercial practice and regulatory realities, co‑developing programmes with growers, greenhouse operators and input manufacturers rather than pushing one‑size‑fits‑all solutions.




Comments